
目次
この記事をシェア
千葉大学大学院園芸学研究院の井川智子准教授らの研究グループ、千葉大学国際高等研究基幹の南川舞准教授、名古屋大学大学院生命農学研究科の榊原均教授、理化学研究所環境資源科学研究センター(理研CSRS)の小嶋美紀子専門技術員らとの共同研究により、植物の発生過程において細胞分裂や形態形成を制御する役割を持つ遺伝子を細胞に導入して発現させることで、培地にホルモンなどの植物成長調節物質(Plant Growth Regulator: PGR)注1)を加えなくても組換え細胞が自発的に分裂・増殖し、器官分化する技術を開発しました。さらに、分化過程における細胞内ホルモン濃度の変化と発現パターンを変化させる遺伝子を明らかにしました。
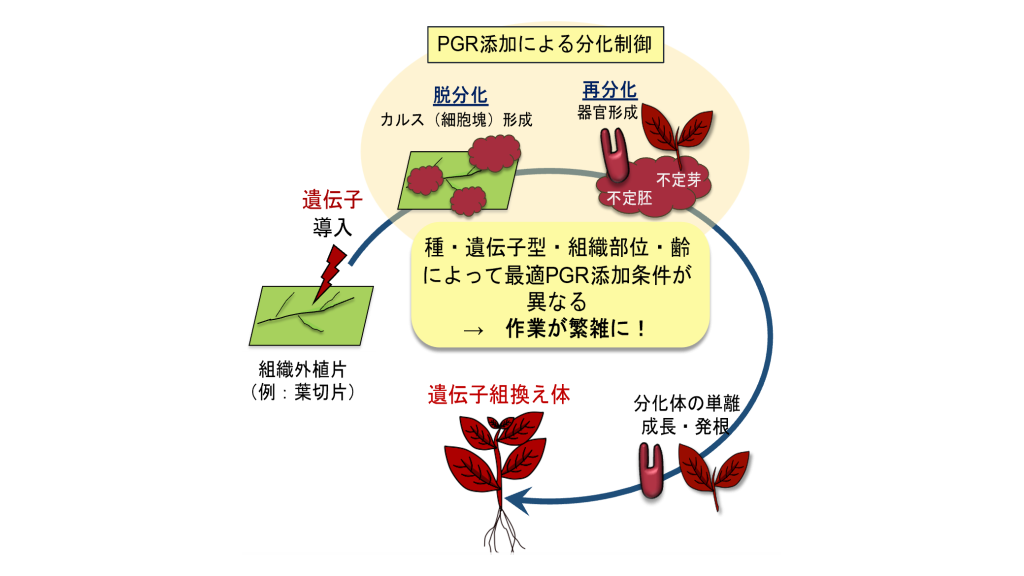
この成果により、植物細胞の分化制御に適切なPGRを添加するための条件を模索する作業を割愛でき、培養による分化制御方法を大きく変革できる可能性を示しています。
本研究成果は、2024年4月3日に国際学術誌Frontiers in Plant Scienceにオンライン公開されました。
■研究の背景
植物では1つの細胞から新しい個体を再生する「分化全能性」が1950年代に実験的に証明されていました。これは培養培地への植物成長調節物質(PGR)添加によって達成されたため、証明から約70年経った現在でも植物細胞の分化制御には培地にPGRを加える方法がとられています。遺伝子導入やゲノム編集といったバイオテクノロジー育種によって改良植物を作製するためには、ゲノムを改変した細胞を分化させて植物体にまで再生させる必要がありますが、分化を導くPGR添加条件は改変対象の植物種ごとに異なるため、バイオテクノロジー育種に取り組む前提として、細胞から個体を分化させる培養技術を個別に構築しておく必要があります。しかし、最適な分化誘導条件を確立するまでには、多くの試験区での検証が必要となり、作業が煩雑となるケースが多い点が現状と課題です(図1)。
近年、植物の発生や形態形成を制御する転写因子注2)(Developmental Regulator: DR)が複数同定され、これらを組織片の体細胞で過剰発現させると従来法よりも分化誘導効率が促進される効果が報告されていました。しかし、培養中の培地へのPGR添加は依然として要求されており、完全なPGRフリー培養条件下での組織培養における分化誘導例はありませんでした。
■研究の成果
本研究ではシロイヌナズナ(Arabidopsis thaliana)由来のBBM(BABYBOOM)注3)とWUS(WUSCHEL)注4)の2種類のDR遺伝子をタバコ(Nicotiana tabacum)の葉片細胞へ導入し,その後PGRフリー培地上で培養して分化誘導における効果を評価しました。解析では、様々な発現制御法およびDR遺伝子の改変を検証しました。その結果、遺伝子を導入しなかった葉片、BBMまたはWUSのみを導入した葉片、改変しないBBMとWUSを導入した葉片は培養4週間後に枯死したのに対して、BBMの転写活性増強型(BBM–VP16)およびWUSを導入した葉片からは、培養4週間後に細胞が脱分化を起こしてカルス注5)が形成されました(図2上段;△)。さらにBBM–VP16とWUS転写抑制増強型の改変遺伝子(SRDX–WUSまたはSRDX–WUSm1)を導入した葉片からは培養2週間後にカルスが発生し、その4週間後には再分化が起こり芽も発生しました(図2下段;⇧)。
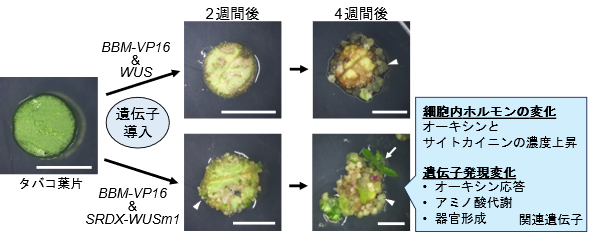
△:カルス(脱分化した細胞の塊)、⇧:再分化した芽
これら分化体であるカルスや芽は組換え細胞から構成され、導入したBBMやWUS遺伝子が発現していることも確認されました。再分化も誘導されたBBM–VP16&SRDX–WUSm1を導入した葉片では、細胞内のオーキシンとサイトカイニンの上昇がより顕著であり、オーキシン応答、アミノ酸代謝、器官形成に関与する遺伝子の発現変動が顕著に検出されました。オーキシンとサイトカイニンは植物の細胞の分化を引き起こす代表的なPGRとして知られており、従来の細胞・組織培養では合成PGRを培地に添加して分化制御に利用されています。本研究では改変型のBBMとWUS遺伝子を共導入した結果、細胞内の天然のオーキシンとサイトカイニン濃度が上昇して、分化反応を導いたと考えられました。そのため、発現が変化した遺伝子中にオーキシン応答性のものが検出された、と考えられます。また、アミノ酸代謝や器官形成に関与する遺伝子の発現が変化していたのは、細胞の分化反応に伴う細胞内の生理や形態の変化を反映していると考えられます。
■今後の展望
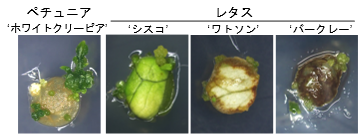
本研究で開発した方法はレタスやペチュニアなどの園芸植物種でも、葉片細胞からの分化を誘導しました(図3)。分化反応自体が遺伝子組換え細胞であることの指標ともなるため、DRと同時に目的遺伝子を導入することで、組換え体を作製しやすくなります。
本成果を基盤として、現在は他植物種での汎用性拡大を目標に、異種間でのBBMおよびWUSの機能保存性の追究や、望みのタイミングで分化を誘導するために厳密な発現制御法の開発を展開させています。
■用語解説
注1)植物成長調節物質 (PGR):植物ホルモンのような分化誘導物質
注2)転写因子:他の遺伝子の発現を調節するタンパク質
注3)BBMタンパク質:植物の受精卵からの胚発生に関与する転写因子
注4)WUSタンパク質:植物の形態形成に関与する転写因子
注5)カルス:器官や組織を構成していた細胞がその個性を失い、分裂を優先させて形成された細胞の塊
■研究プロジェクトについて
本研究は、以下の事業の支援を受けて行われました。
・科学研究費助成事業(JP17K07600,JP19H04852,JP23H04732)
・JST次世代研究者挑戦的研究プログラム
■論文情報
タイトル:Autonomous differentiation of transgenic cells requiring no external hormone application: the endogenous gene expression and phytohormone behaviors.
著者:Sato Y., Minamikawa M.F., Pratama B.B., Koyama S., Kojima M., Takebayashi Y.,Sakakibara H., Igawa T.
雑誌名:Frontiers in Plant Science
DOI:https://doi.org/10.3389/fpls.2024.1308417






